
Гистофизиологические особенности сперматогенеза
Сперматогенез — сложный физиологический процесс образования зрелых мужских половых клеток (ПК) путем редукционного деления и метаморфоза клеток-предшественниц — сперматогоний, протекающий в семенных извитых канальцах яичка.
Яички (семенники)
Парные органы, расположенные в мошонке, основной функцией которых является продукция сперматозоидов и тестостерона. Каждое яичко представляет собой орган овальной формы примерно 4 см длиной, шириной от 2 до 3 см и толщиной 3 см, окруженный плотной соединительнотканной белочной оболочкой, которая утолщается со стороны заднего края и пронизывает паренхиму органа, формируя 200–250 пирамидальных компартментов, известных как дольки (lobuli testis). Каждая долька содержит от 2 до 4 слепо заканчивающихся семенных извитых канальцев, окруженных богато васкуляризированной соединительной тканью, внутри которой распределены конгломераты интерстициальных клеток (Лейдига), ответственные за синтез тестостерона. Сперматозоиды вырабатываются сперматогенным эпителием семенных извитых канальцев, далее следуют в средостение яичка через прямые канальцы, а оттуда — в выносящие канальцы (рис. 1).
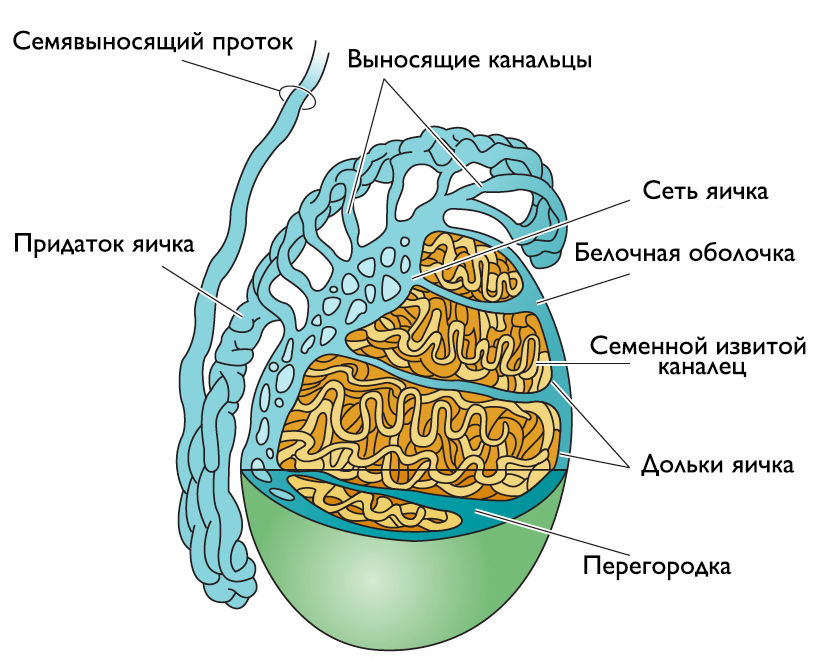
Рисунок 1 | Строение яичка и придатка.
Клетки Сертоли — поддерживающие клетки или сустентоциты
Клетки Сертоли (рис. 2) — высокие столбчатые клетки, плазмалемма которых имеет глубокие цитоплазматические карманы, в которых содержатся ПК на разных стадиях развития. Основанием клетки Сертоли лежат на базальной мембране, а верхушкой обращены в просвет канальцев. На базальной части плазматической мембраны локализованы рецепторы к ФСГ и андрогенам. Латеральные отростки, соединяясь плотными контактами друг с другом, делят семенной каналец на два изолированных компартмента — базальный и адлюминальный. Плотные контакты образуют гемато-тестикулярный барьер, защищающий развивающиеся ПК от иммунной системы.
В основные функции сустентоцитов входят:
- Механическая и трофическая поддержка ПК.
- Фагоцитоз остатков цитоплазмы.
- Формирование гемато-тестикулярного барьера.
- Синтез андроген-связывающего белка (АСБ), который увеличивает концентрацию тестостерона в канальцах путем его связывания и хранения (в 200 раз!).
- Синтез (в процессе эмбриогенеза) антимюллерова гормона, который подавляет образование мюллерова протока (предшественник органов женской половой системы) и направляет развитие эмбриона по мужскому типу.
- Синтез и секреция ингибина — гормона, который ингибирует высвобождение ФСГ передней долей гипофиза.
- Синтез активина — гормона, который стимулирует высвобождение ФСГ из передней доли гипофиза.
- Выделение богатой фруктозой среды, которая питает сперматозоиды и способствует их продвижению к половым путям.
- Синтез и секреция тестикулярного трансферрина — апопротеина, который принимает железо от трансферрина сыворотки и передает его созревающим гаметам.
Процесс сперматогенеза состоит из нескольких этапов превращения сперматогониев в зрелые сперматоциты:
- Сперматоцитогенез — дифференциация сперматогониев в первичные сперматоциты.
- Мейоз — редукционное деление первичных сперматоцитов, благодаря которому уменьшается их хромосомный набор и формируются гаплоидные сперматиды.
- Спермиогенез — трансформация сперматид в сперматозоиды.
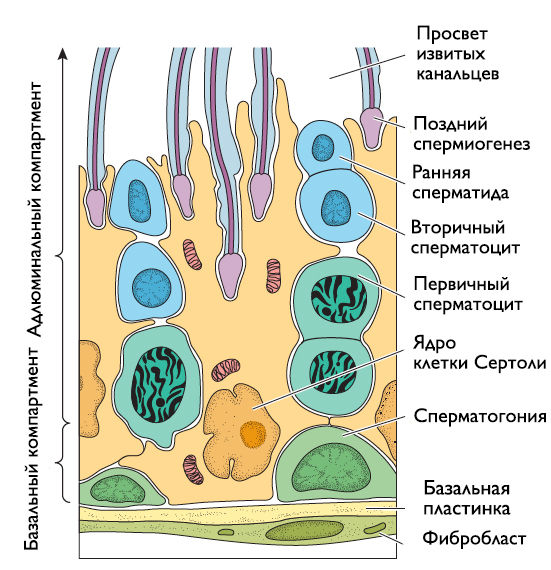
Рисунок 2 | Строение семенных извитых канальцев.
Дифференциация сперматогониев в первичные сперматоциты
Сперматогонии — небольшие диплоидные зародышевые клетки, которые располагаются в базальном компартменте семенных извитых канальцев. Во время полового созревания под влиянием тестостерона покоящиеся ПК включаются в клеточный цикл. Эти клетки бывают следующих типов:
— Темные сперматогонии типа А (истинно стволовые клетки)
Маленькие куполообразные клетки диаметром около 12 мкм с овальным ядром, содержащим большое количество гетерохроматина, который придает ядру темный цвет. Темные клетки служат резервом и не входят в клеточный цикл до стимуляции. Они обладают низкой скоростью митоза и высокой резистентностью к ионизирующей радиации. В случае стимуляции начинают делится митозом и пополняют популяцию сперматогоний типа А.
— Светлые сперматогонии типа А (делящиеся)
Идентичные темным клеткам сперматогонии, отличающееся большим содержанием в эухроматина в ядре. После индукции тестостероном они делятся митозом на две клетки, одна из которых идентична материнской, а другая становится клеткой типа B.
— Сперматогонии типа B
Грушевидные клетки с округлым ядром, делятся митозом и далее дифференцируются в первичные сперматоциты.
Мейоз сперматоцитов
Первичные сперматоциты изначально сходны со сперматогониями типа B, далее они мигрируют из базального в адлюминальный компартмент и почти сразу же вступают в профазу 1 мейоза. Первичные сперматоциты — самые большие клетки сперматогенного эпителия. Они имеют большое везикулярное ядро с конденсированными хромосомами.
Профаза первого мейотического деления длится 22 дня и включает в себя следующие фазы:
- Лептотена — конденсация хромосом с образованием хромосомных нитей.
- Зиготена — конъюгация хромосом с образованием тетрад.
- Пахитена — образование хиазм и кроссинговер.
- Диакинез — максимальная конденсация хромосом, растворение ядерной оболочки и расхождение центриолей.
Обмен гомологичными участками хромосом — кроссинговер — обеспечивает уникальный набор генетического материала каждой гамете и способствует разнообразию человеческого генофонда.
В метафазу 1 парные гомологичные хромосомы выстраиваются вдоль экватора, расходятся по разным полюсам — в анафазу 1. Дочерняя клетка отделяется от материнской в телофазу 1, формируя 2 вторичных сперматоцита.
Вторичные сперматоциты — относительно небольшие клетки, живущие около 10 часов, поэтому их трудно обнаружить с помощью световой микроскопии. Они не удваивают свою ДНК и сразу переходят во второе мейотическое деление, после которого образуются 2 гаплоидные сперматиды. Благодаря модифицированной цитотомии, между гаметами (начиная со сперматогоний типа А и заканчивая сперматидами) остаются цитоплазматические мостики, которые формируют синцитий, позволяющий клеткам синхронизировать свою работу друг с другом и переносить питательные вещества.
Трансформация сперматид (спермиогенез)
Сперматиды — небольшие круглые гаплоидные клетки диаметром 8 мкм, которые, соединяясь друг с другом при помощи цитоплазматических мостиков, образуют небольшие кластеры около просвета канальца. Они имеют большое число митохондрий, хорошо развитый шероховатый ЭПР и комплекс Гольджи. По мере развития сперматиды перестраивают и уменьшают свои органеллы, формируют новые и запасаются гидролитическими ферментами.
Процесс спермиогенеза подразделяется на следуюшие стадии:
Стадия формирования акросомального пузырька (рис. 3)
Сначала гидролитические ферменты образуются на мембранах шероховатой ЭПР, далее модифицируются в комплексе Гольджи и затем упаковываются транс-сетью Гольджи в маленькие мембраносвязаные преакросомальные гранулы. Гранулы начинают сливаться и образуют акросомальный пузырек, который подходит к передней поверхности ядерной оболочки и связывается с ней. Акросомальный пузырек продолжает расти в размере и распластывается по передней поверхности ядра, образуя “шапочку”.
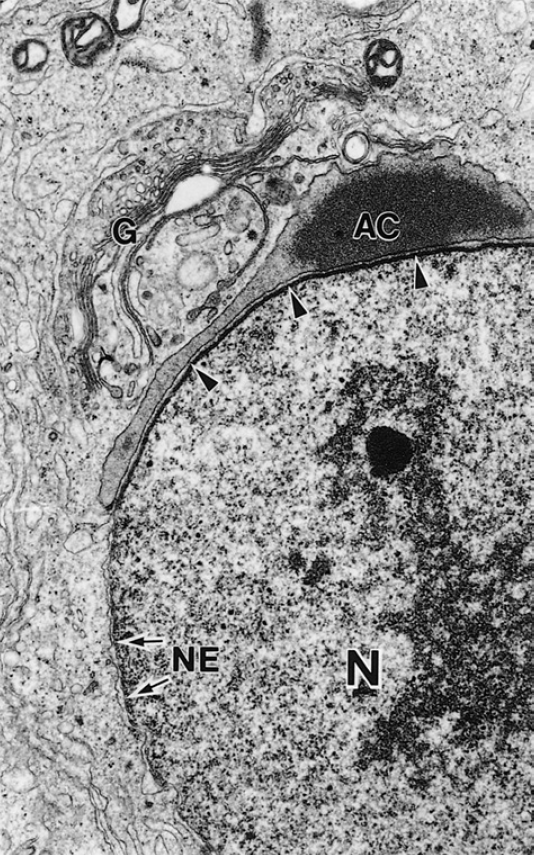
Рисунок 3 | Электронная микрофотография стадии формирования акросомального пузырька (Å~18000). AC — акросома; G — аппарат Гольджи; N — ядро; NE — ядерная мембрана.
Стадия формирования хвоста
Ядро уменьшается в размерах (за счет уплотнения хромосом) и уплощается, принимая характерную форму. Микротрубочки одной из центриолей собираются в промежуточную структуру — манжету, которая, удлиняясь, образует жгутик, а затем исчезает. Цитоплазма окружает основание хвоста, и митохондрии образуют цилиндр вокруг аксонемы.
Стадия созревания (рис. 4)
Характеризуется уменьшением объема цитоплазмы путем фагоцитоза клетками Сертоли, после этого синцитиальные связи разрушаются, и самостоятельные сперматозоиды высвобождаются в просвет извитых канальцев. Стоит отметить, что вновь образованные сперматозоиды неподвижны и не способны к оплодотворению, они приобретают подвижность только после прохождения каналов придатка яичка.
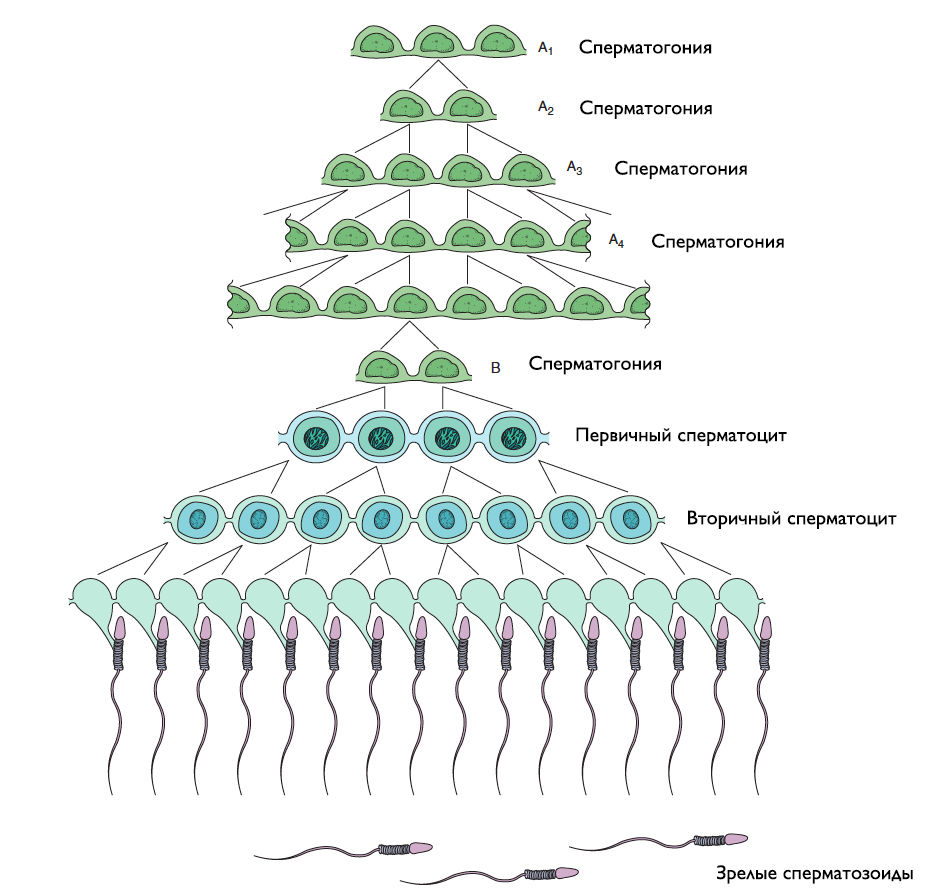
Рисунок 4 | Сперматогенез и межклеточные связи синцития при дифференцировке и созревании.
Морфологическое строение сперматозоида
Сперматозоиды (рис. 5) — длинные клетки (65 мкм), образующиеся в процессе сперматогенеза. Каждый сперматозоид состоит из головки, шейки и хвоста.
Уплощенная головка длиной в 5 мкм содержит ядро с конденсированным хроматином и акросому. Акросома располагается кпереди от ядра и содержит разнообразные ферменты: нейраминидазу, гиалуронидазу, кислую фосфатазу, арилсульфатазу и трипсиноподобную протеазу, известную как акрозин. Связывание сперматозоида с гликопротеином блестящей оболочки ZP3 запускает каскад реакций, способствующих оплодотворению.
Хвост подразделяется на 4 отдела: шейку, промежуточный, главный и дистальный. В основании хвоста в виде оси располагается аксонема, представленная микротрубочками, которые описываются формулой 9 х 2 + 2, т. е. 9 дуплетов микротрубочек по периферии и один дуплет в центре. В дистальном отделе хвоста других структур нет, кроме аксонемы, распадающейся в конце на отдельные микротрубочки. В главной и промежуточной части хвоста вокруг аксонемы располагаются 9 продольно ориентированных плотных одиночных элементов цитоскелета — миофибрилл, формирующих волокнистый футляр осевой нити. В промежуточной части вокруг фибрилл, окружающих аксонемы, находятся митохондрии, образующие митохондриальную спираль.
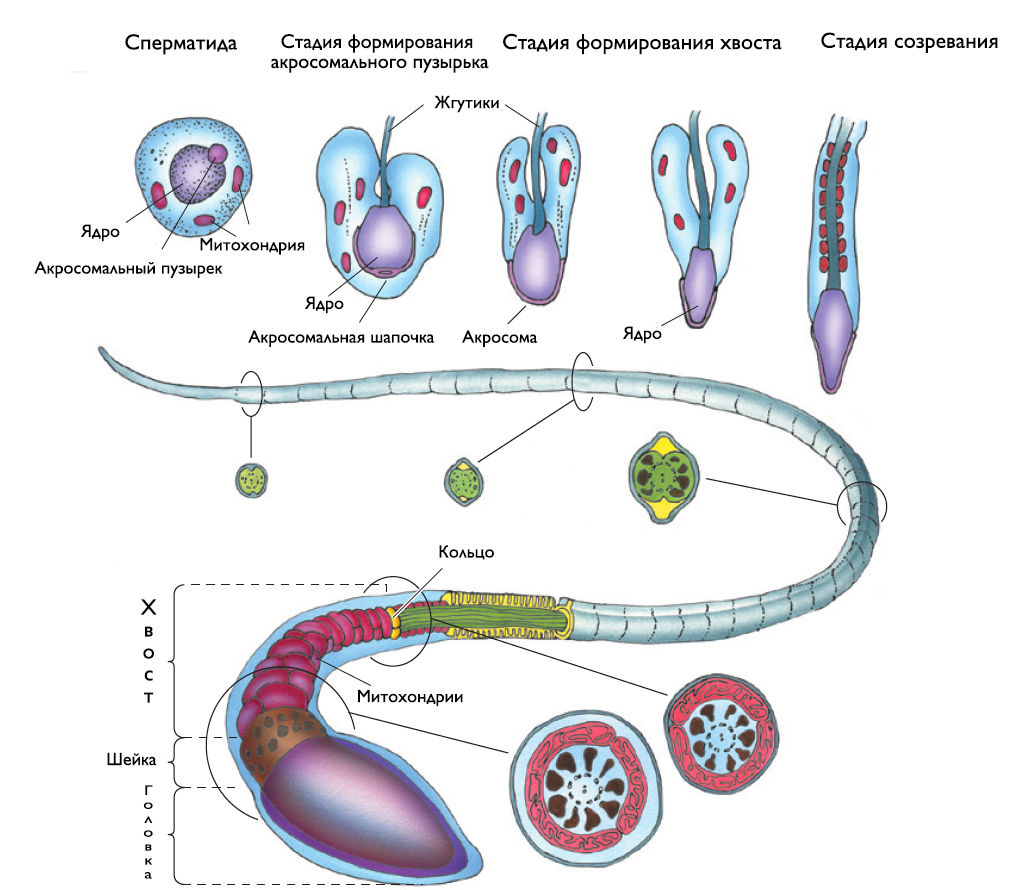
Рисунок 5 | Строение сперматозоида.
Цикличность сперматогенеза
Известно, что все половые клетки на разных этапах сперматогенеза соединены между собой синцитиальным мостиками, что дает им способность без труда взаимодействовать друг с другом и синхронизировать свою деятельность. Тщательное исследование сперматогенного эпителия позволило выявить шесть типов одинаковых клеточных ассоциаций на протяжении всего извитого канальца, которые именуются шестью стадиями сперматогенеза (рис. 6). Расстояние между двумя одинаковыми стадиями называется волной сперматогенеза. Таким образом, у человека шесть повторяющихся волн, соответствующих шести стадиям. Общая продолжительность сперматогенеза составляет около 64 дней, но сперма вырабатывается непрерывно, благодаря тому, что разные участки семенных извитых канальцев находятся на разных стадиях.
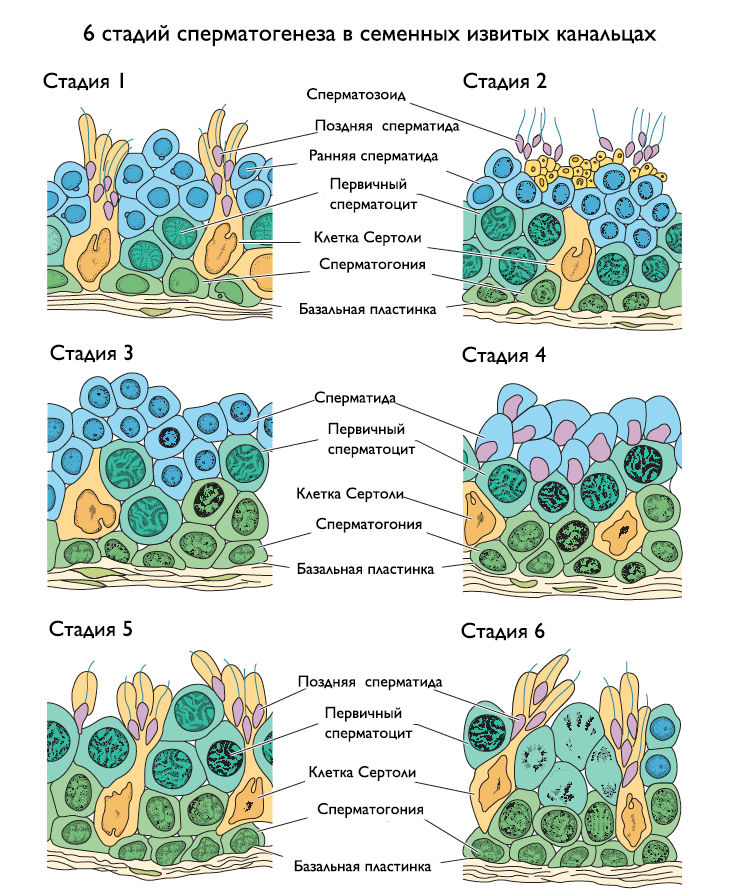
Рисунок 6 | Шесть стадий сперматогенеза, протекающих в семенных извитых канальцах.
Интерстициальные клетки Лейдига
Семенные канальцы окружены богато васкуляризированной рыхлой волокнистой соединительной тканью, которая, помимо своих классических компонентов (фибробласты, тучные клетки и др.), содержит скопления клеток Лейдига. Основная функция этих клеток — продукция тестостерона и инсулиноподобного фактора 3 (INSL 3). Первый отвечает за нормальный сперматогенез, второй — за рост и развитие направляющей яичко связки, и, следовательно, за опускание яичка в мошонку. Интерстициальные клетки — клетки округлой или полигональной формы с одним ядром, в диаметре составляют около 15 мкм. Как типичные стероид-продуцирующие клетки, они имеют митохондрии, хорошо развитую гладкую ЭПС и аппарат Гольджи. Ацидофильная цитоплазма содержит многочисленные пероксисомы, лизосомы, гранулы липофусцина, липидные капли, а также кристаллы Рейнеке, функция которых до сих пор неясна.
Гистофизиология сперматогенеза
Оба яичка за сутки образуют около 120 миллионов сперматозоидов. Клетки Сертоли производят богатую фруктозой жидкость, которая питает и транспортирует молодые сперматозоиды из просвета канальцев в экстратестикулярные семявыносящие пути. Лютеинизирующий гормон, выделяющийся из передней доли гипофиза, связывается с ЛГ-рецепторами клеток Лейдига и запускает аденилатциклазный каскад, что приводит к образованию цАМФ. Активированная цАМФ протеинкиназа индуцирует холестерол-эстеразу, которая отщепляет холестерин от липидных капель. Далее холестерин проходит ряд сложных ЛГ-зависимых химических превращений, которые протекают в митохондриях и эндоплазматическом ретикулуме, откуда в итоге высвобождается тестостерон (рис. 7). Уровень ЛГ контролируется по механизму отрицательной обратной связи тестостероном и дигидротестостероном, тогда как уровень ФСГ — активином и ингибином (рис. 8). Нормального уровня тестостерона плазмы крови недостаточно для сперматогенеза. Поэтому существует андроген-связывающий белок, который связывает тестостерон плазмы и удерживает его в канальцах, обеспечивая его достаточную концентрацию в окружающем пространстве сперматоцитов. Помимо этого, тестостерон необходим для нормального функционирования семенных пузырьков, простаты и бульбоуретральных желез, а также для развития вторичных мужских половых признаков. Клетки организма, нуждающиеся в тестостероне, обладают ферментом 5α-редуктазой, который превращает тестостерон в более активную форму — дигидротестостерон.
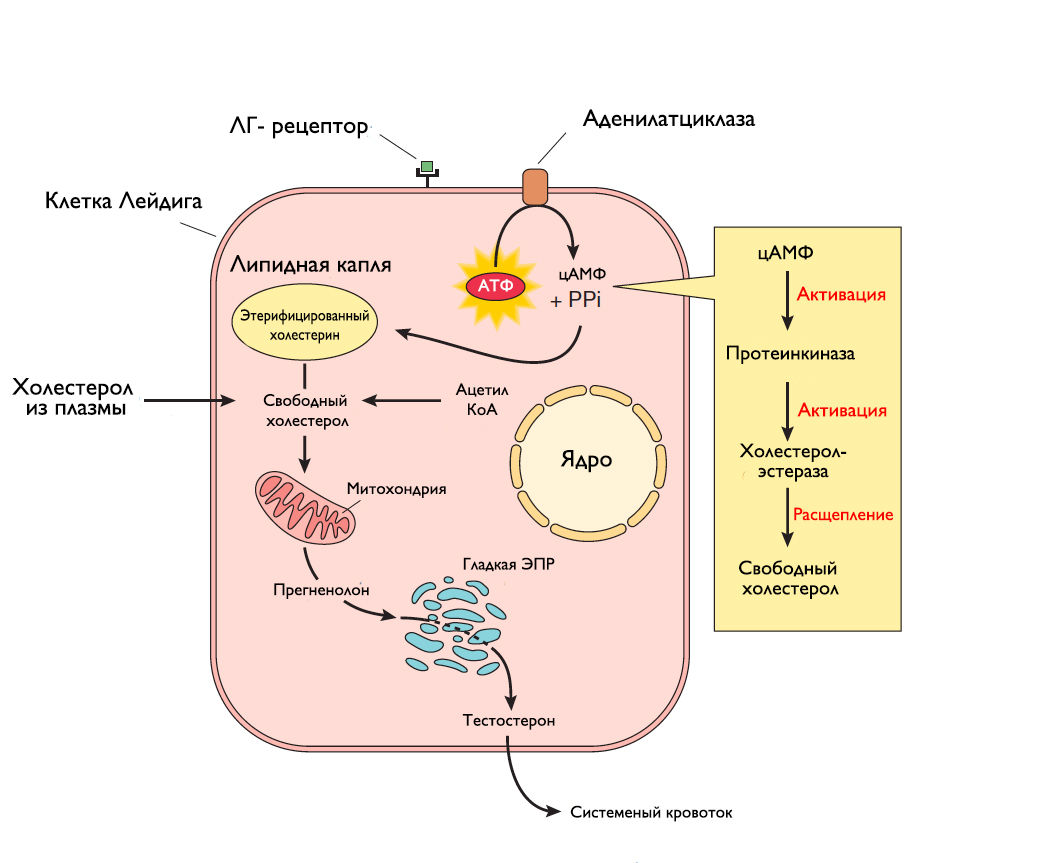
Рисунок 7 | Синтез тестостерона клетками Лейдига.
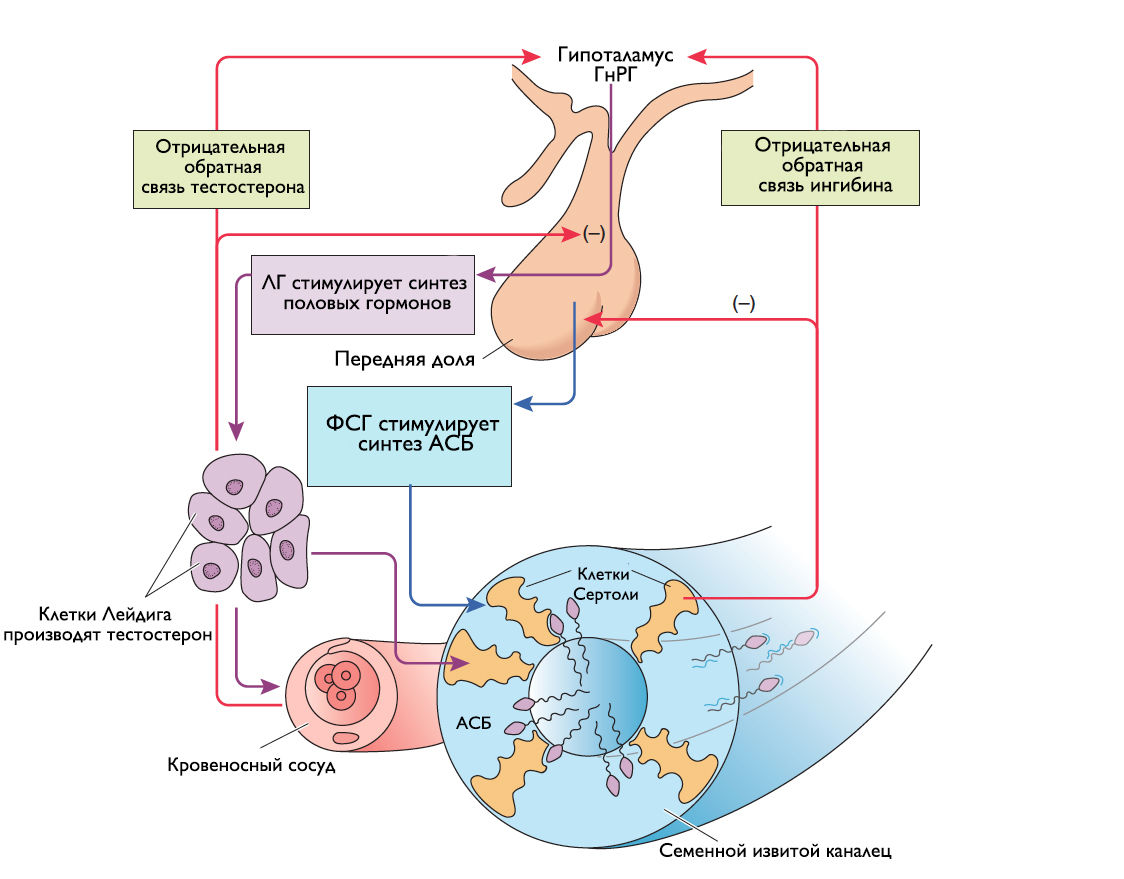
Рисунок 8 | Гормональный контроль сперматогенеза.
Источники:
- TEXTBOOK OF HISTOLOGY, FOURTH EDITION, Leslie P. Gartner, PhD Professor of Anatomy (Ret.), все рисунки взяты отсюда же.
- The Cycle of the Seminiferous Review Epithelium in Humans: A Need to Revisit? RUPERT P. AMANN
- Spermatogenesis and Cycle of the Seminiferous Epithelium. Rex A. Hess and Luiz Renato de Franca.
- Endocrine regulation of sperm release. Kumar, S. Raut and N. H. Balasinor.
